一文读懂三维基因组
每个人体内都有着两米长的DNA,它是如何紧密折叠在直径 10 微米小的细胞核内。
而且还要在极度压缩的情况下,精确地调度数量庞大的调控元件去表达两万多个基因。
如果其中一些关键环节出现问题,随之而来的就是遗传疾病,甚至癌症。
接下来,我们进入细胞核的世界,从三维空间的角度,看看它是如何组织,协调这么庞大的工程。
一、细胞核 Nucleus
首先看下染色质的主要存储容器——细胞核,主要看与三维基因组密切相关的三个结构:
核膜 Nuclear Envelope:用来包裹染色质,控制物质进出
**核纤层 Nuclear Lamina:**位于核膜的内表面的纤维网络,支持核膜,锚定染色质,与核骨架相连,参与细胞周期解离和重建
*核仁 Nucleolus:* 主要存储合成 rRNA,存储装配核糖体
二、染色质疆域 Chromosome Territory,CT
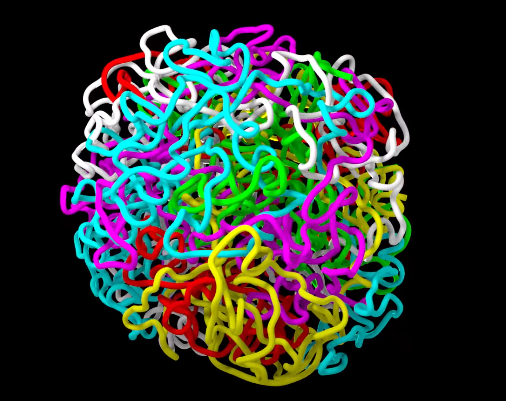
在真核生物的基因组中,细胞核内的染色质分布并不是随机的,为了跨越较大的基因组距离去互相作用,比如增强子和启动子的互作,这些密切接触的染色质会靠的更近,这就是染色质疆域。大概就像这样:

我们可以使用染色体构象捕获技术(3C,4C,Hi-C,HiChIP)来获取到3D基因组。
在二维视角下的染色质疆域
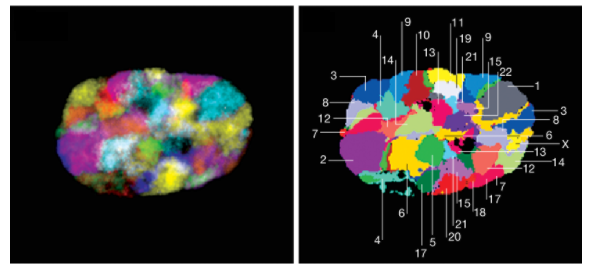
在三维视角下的染色质疆域
目前,发现这些区域有一定的规律:
- 染色体的位置相对不变:这种相对不变会持续到有丝分裂开始。比如大型的,基因贫乏的染色体通常位于核层附近的外围,而较小的,富含基因的染色体则更靠近核中心。
- 染色质的位置会因细胞类型不同而改变:例如,X染色体已显示在肝细胞中比在肾细胞中更频繁地定位在外围
- 同源染色体在细胞间期倾向于彼此分离
为了更方便的研究,进一步把这些互作部分划分为:
三、染色质区室 A/B compartments
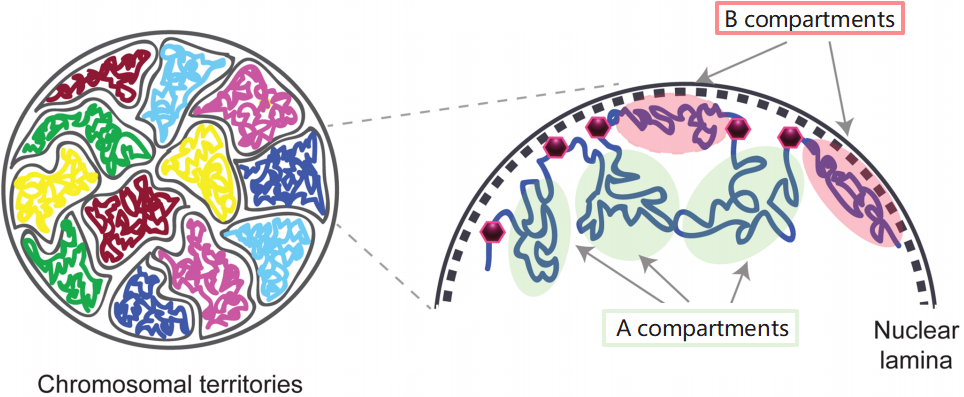
使用 Hi-C 发现,整个基因组被分割为两个空间区室,分布标记为 A,B 染色质区,往往区室内互作频繁,而区室间互作较少。
A compartments:开放的染色质,表达活跃,基因丰富,具有较高的GC含量,包含用于主动转录的组蛋白标记,通常位于细胞核的内部。
B compartments:关闭的染色质,表达不活跃,基因缺乏,结构紧凑,含有基因沉默的组蛋白标志物,位于核的外围。它们主要由LAD组成,包含晚期复制起点。
在生物信息分析中,我们通过计算染色体内部互作的相关性来区分两种不同的区室。

四、拓扑结构域 Topologically associating Domains,TAD
在染色质区室中,我们还会发现互相作用相对频繁的基因组区域,这些就是拓扑结构域 TAD。
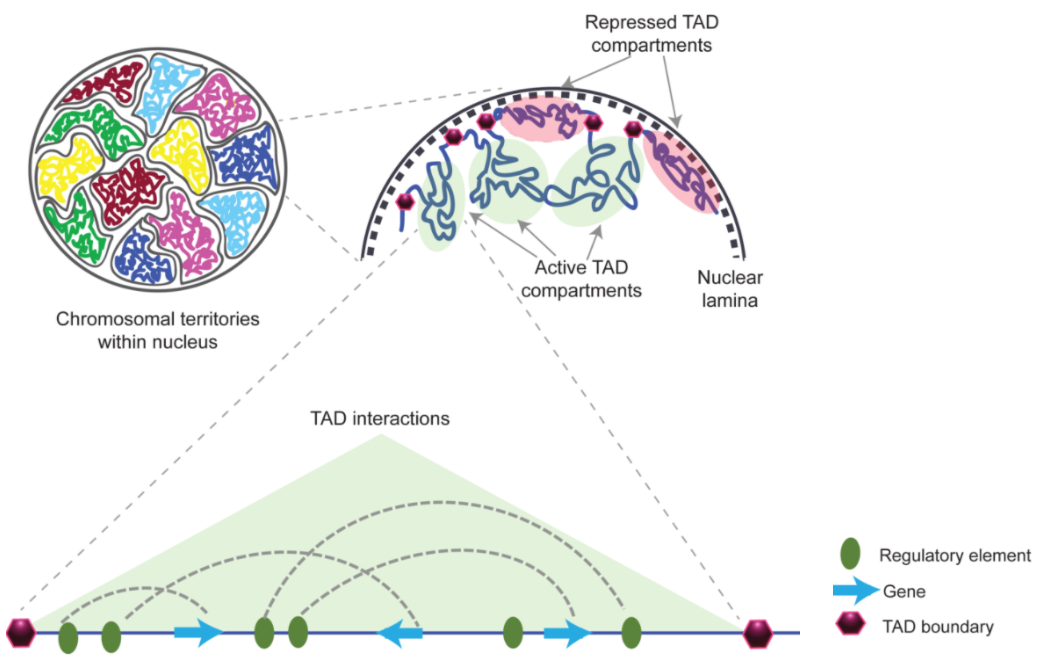
一般这些区域在不同的哺乳动物的不同细胞中都很保守,并且高度富集 CTCF 和 粘附蛋白。
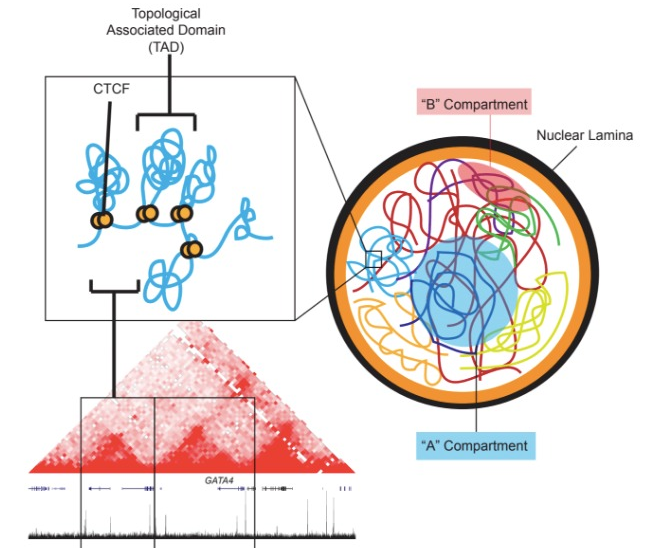
通过计算基因互作矩阵,我们会得到一个类似上图的大三角形,可以看到上面有几个深红色的三角,代表内部高度互作域被定义为拓扑结构域,一般是400-800kb较稳定的复制单元。
TAD 的边界:
- 通过与上下游的互作频率趋近于0的DI值界定TAD边界
- 边界中富含绝缘子蛋白 CTCF 结合位点、活跃转录标记,如H3K4me3及H3K36me3
- 富含结构蛋白结合位点,与管家基因,转运RNA基因和短间隔核元素(SINEs)相关的表观遗传标记。
目前研究最多的是,TAD通过限制每个TAD的增强子-启动子相互作用来调节基因表达,但是TAD详细功能还有待发现。
尽管许多蛋白质复合物,DNA 元件与TAD边界相关,但TAD形成的基础机制也很复杂,尚未完全阐明。
目前认可的模式是,以CTCF蛋白为核心,在黏附蛋白的帮助下,通过loop extrusion模型挤压形成染色质环,锚定TAD边界,为TAD的形成提供了结构基础。此外,TAD 边界的刚度本身可能会导致 TAD 的形成。
TAD 可细分为 sub TAD, 大约长 100kb,sub TAD之间的边界在不同细胞组织间具有差异,与细胞特异性的增强子-启动子互作有关。
在细菌中,这种互作结构叫做染色质互相作用域(Chromosomal Interacting Domains,CIDs)
五、层关联域 Lamina Associating Domains,LAD
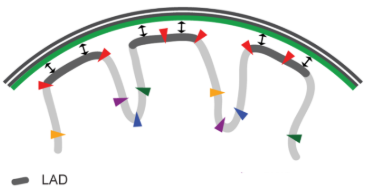
LAD约占基因组的40%,大小介于40kb至30Mb之间,基因较少。 LAD主要由转录沉默染色质组成,富含组蛋白H3K27me3 ,这是异染色质的常见翻译后组蛋白修饰。
结构性 LAD,constitutive LAD,cLAD:富含AT的异染色质区域,靠近在核纤层上,这些区域对染色体之间的结构形成至关重要。
兼性 LAD, facultative LAD,fLAD:具有不同的核纤层相互作用,在不同细胞中包含不同的被激活或抑制基因,从而导致不同的细胞类型。
六、核仁关联域 Nucleolar Associating Domains,NAD
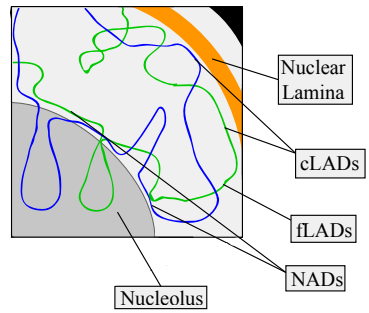
NAD占基因组的4%,几乎具有与LAD相同的所有物理特征。通过对LAD和NAD的序列分析发现,某些区域可能在核纤层和核仁间切换。
七、染色质环 Chromatin loops
染色质在空间中形成环状结构,因此相距很远的染色质区域也可以在三维空间中聚集在一起。
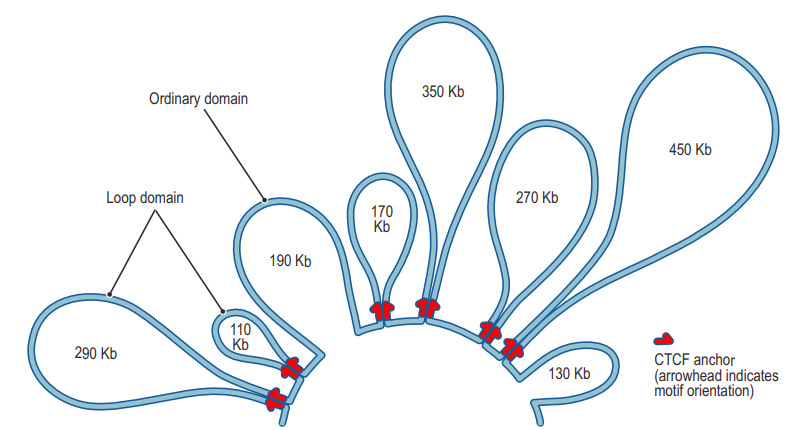
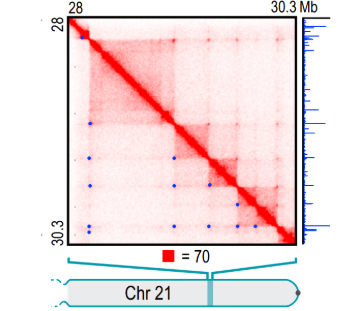
据推测大约50%的人类基因通过染色质环化过程参与长距离的染色质相互作用。我们可以基于基因互作矩阵,来查看互作频率相对周围较强的区域,在下图中用蓝色圆圈标记,这些位置就是为染色质环区域。
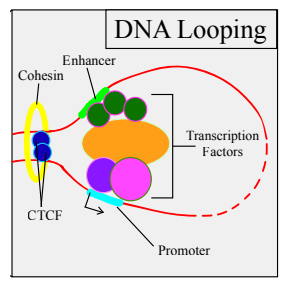
这种结构可以使在线性距离很远的元件得以相遇,以此来调控生命活动,比如,从空间上拉近启动子和增强子的距离,促使基因的转录起始。
这个过程中,接触到启动子的增强子元件可以募集大量蛋白质复合物,例如介导复合物,PIC和细胞特异性转录因子。另外,许多因素也会促进该过程,包括结构蛋白(主要是CTCF和Cohesin),共激活因子和ncRNA等。
下一篇,我们来详细看看三维基因组检测技术:染色体构象捕获技术(3C,4C,Hi-C,HiChIP)
参考
The new cytogenetics: blurring the boundaries with molecular biology
Regulation of disease-associated gene expression in the 3D genome
Chromatin Domains: The Unit of Chromosome Organization
Minor Loops in Major Folds: Enhancer–Promoter Looping, Chromatin Restructuring, and Their Association with Transcriptional Regulation and Disease
Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome
A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping
The Three-Dimensional Organization of Mammalian Genomes
Enhancer activation requires trans-recruitment of a mega transcription factor complex
https://foo-lab.com/services/chromatin-architecture-epigenetics/
 微信
微信 支付宝
支付宝








